Heinz-Dieter Hoppe
Begriffsbestimmung
Im Allgemeinen wird mit dem Begriff „Landsorte“ eine durch den Menschen kultivierte, genetisch heterogene Sorte definiert. Sie hat sich in einem bestimmten ökogeografischen Gebiet über Jahrhunderte entwickelt und an die dortigen boden- und klimatischen Bedingungen angepasst. Sie ist immer an die jeweilige traditionelle Bewirtschaftung und Nutzung gebunden. Da sich die Umwelt jährlich ändert, passt sie sich kontinuierlich an. Sie befindet sich in einem ständigen Evolutionsstadium. Eine Landsorte besitzt die Fähigkeit, biotischen und abiotischen Stress zu tolerieren, was zu einer hohen Ertragsstabilität und einem mittleren Ertragsniveau in einem landwirtschaftlichen System mit geringem Input führt (Zeven 1998).
Landsorten werden auch als einheimische Sorten, traditionelle Sorten, historische Sorten, lokale Sorten, Heirloom-Sorten, Hausgarten-Sorten oder Bauernsorten bezeichnet.
Die Domestizierung von Wildarten durch den Menschen führte durch natürliche und unbewusste/bewusste Selektion zur Einengung der genetischen Vielfalt der Ausgangspopulationen. Daraus entstandene Landsorten besitzen im Regelfall immer noch eine höhere genetische Variabilität als Zuchtsorten (Tanksley und McCouch 1997; Dias et al. 2024) (Abb. 1).
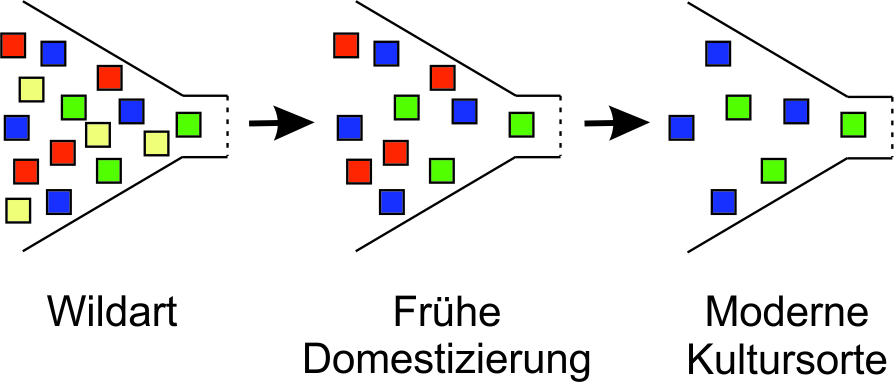
Abb. 1. Genetische Engpässe (Flaschenhalseffekt), die Nutzpflanzen während der Domestizierung und durch moderne Pflanzenzüchtungspraktiken auferlegt wurden. Die Kästchen stellen Allel-Variationen von Genen dar, die ursprünglich in Wildarten existierten, aber durch Domestizierung und Züchtung allmählich verloren gingen. Grafik modifiziert nach Tanksley und McCouch (1997)
Der Begriff „Landsorte“ wird seit Ende des 19. Jahrhunderts in wissenschaftlichen Kreisen diskutiert. Es wurde damals bereits erkannt, dass es sich dabei um wertvolle genetische Ressourcen handelt. Um diese zu erhalten, erfolgte die Einrichtung von Genbanken. Das Konzept einer Genbank wurde erstmalig vom Namensgeber des heutigen N. I. Vavilov Instituts für Pflanzengenetische Ressourcen (VIR) in St. Petersburg realisiert (Lehmann 1981).
A.C. Zeven von der Landwirtschaftlichen Universität Wageningen (Niederlande) beschäftigte sich intensiv mit dem Thema „Landsorte“. Er kam zu dem Schluss, dass Landsorten eine recht komplexe Natur aufweisen und es nicht möglich ist, eine allumfassende Definition zu geben, da dies lediglich zu einer Beschreibung führen würde (Zeven 1998). Diese Schlussfolgerung trifft den Kern, da die Entwicklung der Landsorten eng mit natürlicher Selektion und/oder menschlichem Handeln verwoben ist.
Neuartige molekulargenetische Verfahren, wie AFLP-Analyse oder SNP-Genotypisierung, gestatten mittlerweile die gezielte Untersuchung von der in Landsorten enthaltenen genetischen Vielfalt und deren Beziehung zu modernen Sorten.
An Hand von drei ausgewählten Beispielen wichtiger europäischer Kulturpflanzenarten soll dargestellt werden, dass auch unter den „Landsorten“ Formen mit geringerer genetischer Diversität anzutreffen sind. Gleichwohl stellen Formen dieser Gruppe eines der wichtigsten Reservoire für die Nutzung und Erhaltung von genetischer und morphologischer Diversität dar.
Roggen (Secale cereale)
Roggen ist mit Weizen und Gerste als Beikraut eingewandert und hat seine Karriere auf leichten, wasserfernen Böden Deutschlands sowie Nord- und Osteuropas angetreten. Im Mittelalter erfolgte eine Selektion dadurch, dass die Bauern Ähren und Stroh separat ernteten, um den Anteil von Unkrautsamen im Erntegut zu reduzieren (Zeven 2000). Wahrscheinlich nutzten die Bauern nur die größten und attraktivsten Ähren für die nächste Aussaat. Diese Art der Selektion stellt keine strenge Selektion dar. Phänotypen, die an besser mit Nährstoffen und Wasser versorgten Stellen aufwuchsen und damit einen Vorteil bei der Ährenbildung hatten, wurden mit genetisch ertragreichen Phänotypen vermischt. So ist davon auszugehen, dass den größten Einfluss auf diese Populationen die natürliche Evolution hatte.
Untersuchungen von Persson und Bothmer (2000) an schwedischen Roggenlandsorten zeigten, dass die genetische Diversität von Landsorten in etwa der von um 1990 in Schweden angebauten Zuchtsorten entsprach. Diese Aussage wiederspricht allerdings der allgemeinen Annahme, dass Landsorten per se eine höhere genetische Variabilität im Vergleich zu modernen Sorten aufweisen (Dias et al. 2024). Die Autoren der Studie vermuten, dass auf Grund zu kleiner Populationen eine genetische Drift eintrat. Hier wird deutlich, dass gerade bei fremdbefruchtenden Arten die Anzahl der Samenträger pro Bestäubungsfläche von entscheidender Bedeutung für den Erhalt der genetischen Variabilität ist (Zeven 2002)!
Tomate (Solanum lycopersicum)
Die Tomate ist in Deutschland das beliebteste Gemüse und weltweit die wichtigste Gemüseart. Im Gegensatz zu Roggen ist diese Art nicht natürlich eingewandert, sondern vor ca. 500 Jahren durch den Menschen als bereits bestehende Kulturpflanze nach Europa eingeführt. Damalige Transportmöglichkeiten und die Geschwindigkeit im Seeverkehr waren nicht unbedingt dafür geeignet, empfindliches Pflanzenmaterial, wie z.B. Früchte, unversehrt von Mittelamerika nach Spanien zu bringen. Es ist anzunehmen, dass Saatgut nur von einzelnen Pflanzen, schon gar nicht von Populationen, nach Europa gelangte. Das von diesen Pflanzen abstammende Saatgut bildete die Grundlage für die heutige Tomatenvielfalt auf unserem Kontinent. Nach Untersuchungen von Miller und Tanksley (1990) an Wildformen, lateinamerikanischen Landsorten sowie amerikanischen Zuchtsorten befindet sich in der Kulturform Solanum lycopersicum, wie wir sie kennen, nur noch ein Bruchteil der möglichen genetischen Vielfalt der Sektion Lycopersicon (Abb. 2).

Abb. 2. Genetische Vielfalt der Kulturtomate (gelb) innerhalb der Sektion Lycopersicon. Grafik nach Miller und Tanksley (1990)
Die wahrscheinlich bisher umfangreichste Untersuchung von traditionellen Tomaten-Akzessionen führten Pons et al. (2022) durch. Sie analysierten 1.499 Herkünfte aus dem Mittelmeerraum (TRADITOM-Sammlung – EU Funding & Tenders Portal (2025)). Parallel zur genotypischen Klassifizierung wurden alle Akzessionen in fünf Ländern an insgesamt zehn Standorten phänotypisch charakterisiert. Die Ergebnisse zeigen eine beeindruckende Variabilität in den Fruchteigenschaften, die für diesen Genpool einzigartig ist.
So sind Mutationen für langfrüchtige und lagerfeste Formen nur in Europa zu finden. In den Ursprungspopulationen Latein- und Mittelamerikas treten sie nicht auf (Rodríguez et al. 2011).
Zu den langfrüchtigen Formen Europas zählt z.B. der San Marzano-Typ.
Landsorten, die unter Trockenstress und geringem Input Früchte produzieren, die für eine Langzeitlagerung (drei bis sechs Monate) geeignet sind, wurden unabhängig voneinander in Spanien und Italien entwickelt. Diese Akzessionen werden LSL-Tomaten (LSL = Long-Shelf-Life) genannt.
Um der großen Nachfrage nach lagerfesten Sorten gerecht zu werden, entstand die Idee, LSL-Tomaten zusätzlich im Gewächshaus anzubauen. Figàs et al. (2018) prüften 12 Sorten (sieben Landsorten, drei Selektionen und zwei Hybriden) parallel im Freiland (FL) und Gewächshaus (GWH). Die Ergebnisse waren ernüchternd. Die Früchte der Pflanzen aus dem GWH hatten ein geringeres Gewicht, ihre Festigkeit war reduziert und optisch waren sie weniger rot als solche aus dem FL. Im Durchschnitt war der Ertrag bei den GWH-Pflanzen um 35 % niedriger und der tägliche Gewichtsverlust der Früchte nach der Ernte um 41 % höher als nach FL-Anbau. Diese Ergebnisse zeigen eindringlich, wie weit hier durch die Selektion der Bauern eine ganz enge Anpassung an lokale Umweltbedingungen erfolgt ist.
Gemüsekohl (Brassica oleracea)
Die große Vielfalt an unterschiedlichen Kohlformen wäre ohne den Einfluss des Menschen nicht möglich gewesen. Die Natur interessiert sich nicht für unterschiedliche Nutzungsarten, wie Kopfkohl, Blattkohl oder Kohlrabi, es geht nur um das Überleben der Art.
In Süditalien und auf Sizilien entwickelten Bauern unterschiedlich genutzte Landsorten. Stansell und Björkman (2020) untersuchten 109 Brokkoli- oder Brokkoli-/Blumenkohl-Zwischenformen dieser Regionen mittels Genotypisierungsmarker. Die gesammelten Landsorten zeigten pro Akzession eine wesentlich höhere genetische Diversität als F1-Hybriden. Das war zu erwarten.
Wie sieht es dagegen bei ewigem Grünkohl aus? Viele der Formen sind reine Klone, die die Fähigkeit zu blühen verloren haben und daher über bewurzelte Zweige vermehrt werden müssen (Zeven 2000). Von Diversität kann hier nicht mehr geredet werden, es liegt nur noch ein Genotyp vor.
Die wenigen angeführten Beispiele zeigen, wie weit der Begriff „Landsorte“ gefasst ist. Landsorten und Zuchtsorten sind Produkte der menschlichen Schöpferkraft. Sie können mit ihren Gebrauchseigenschaften auch nur in einer durch Menschen geprägten (regulierten) Umwelt existieren. Bei einem Anbau von Kulturpflanzen mit freier Abblüte und ohne Selektion würden diese sich nach mehreren Generationen wieder in Richtung Wildtyp entwickeln oder völlig verschwinden. Die überlebenden Akzessionen hätten dann mit Sicherheit eine höhere Klimaresilienz. Ob das dann in jedem Fall zielführend für die Ernährungssicherheit und Essgewohnheiten der Menschen wäre, sei dahingestellt.
Entscheidend ist, wie wir mit den uns zur Verfügung stehenden Möglichkeiten die noch existierende Diversität erhalten, ohne Gebrauchswerteigenschaften zu verlieren.
Erhalterorganisationen wie der VERN, der VEN oder ARCHE NOAH praktizieren seit Jahrzehnten die On-farm-Erhaltung. Eine der wichtigsten Möglichkeiten um Kulturpflanzen zu erhalten und weiter zu entwickeln.
Von staatlicher Seite erfolgt allerdings nur eine rudimentäre Unterstützung dieser vielfach ehrenamtlichen Arbeit, es werden im wesentlichen Genbanken favorisiert. So wichtig wie sie sind, findet durch die sehr langen Vermehrungszyklen (erneute Vermehrung zum Teil erst nach über 30 Jahren (Börner et al. 2008)) kein lebendiger Erhalt von Vielfalt statt. Ferner kann es auch in Genbanken zu Verlusten an Allelen führen, da bei kritischen Wetterereignissen (z.B. Auswinterung) nicht alle Pflanzen überleben und die Vermehrungsflächen zu klein sind, um diesen Selektionsdruck zu kompensieren (Bolibok-Brągoszewska et al. 2014; Chebotar et al. 2003).
Parallel zu Genbanken sollte die On-farm-Erhaltung wesentlich stärker gefördert werden. Dabei sind neben Erhalterorganisationen auch gärtnerische und landwirtschaftliche Betriebe einzubeziehen. Gerade bei Letzteren können ausreichend große Populationen etabliert werden, die eine genetische Drift verhindern. Das gelingt allerdings nicht mit der bisherigen Förderpraxis von kurzzeitigen Projekten. Die Betriebe müssen langfristig in die Lage versetzt werden, auskömmlich mit Landsorten zu wirtschaften. Der Effekt wäre nicht nur der Erhalt von genetischer Vielfalt, sondern durch die notwendige Low-Input-Bewirtschaftung könnte z.B. der Humuserosion von Böden entgegengewirkt werden. Weitere positive Effekte auf die Agrar-Ökosysteme sollen jetzt nicht weiter besprochen werden.
Durch koordinierte Absprachen zwischen Wissenschaft, Erhalterorganisationen und anderen gesellschaftlichen Akteuren könnte die Erhaltung der genetischen Diversität in einem größeren Umfang als Citizen-Science-Projekt durchgeführt werden.
Literaturverzeichnis
Bolibok-Brągoszewska, H.; Targońska, M.; Bolibok, L.; Kilian, A.; Rakoczy-Trojanowska, M. (2014): Genome-wide characterization of genetic diversity and population structure in Secale. In: BMC Plant Biology 14, S. 184. DOI: 10.1186/1471-2229-14-184.
Börner, A.; Nagel, M.; Landjeva, S., Lohwasser, U. (2008): Genetische Diversität in ex situ Genbanken – Erhalt und Nutzbarmachung. In: Vorträge für Pflanzenzüchtung 77, S. 159–161.
Chebotar, S.; Röder, M. S.; Korzun, V.; Saal, B.; Weber, W. E.; Börner, A. (2003): Molecular studies on genetic integrity of open-pollinating species rye (Secale cereale L.) after long-term genebank maintenance. In: TAG. Theoretical and applied genetics. Theoretische und angewandte Genetik 107 (8), S. 1469–1476. DOI: 10.1007/s00122-003-1366-1.
Figàs, M. R.; Prohens, J.; Raigón, M. D.; Pereira-Dias, L.; Casanova, C.; García-Martínez, M. D. et al. (2018): Insights Into the Adaptation to Greenhouse Cultivation of the Traditional Mediterranean Long Shelf-Life Tomato Carrying the alc Mutation: A Multi-Trait Comparison of Landraces, Selections, and Hybrids in Open Field and Greenhouse. In: Frontiers in plant science 9, S. 1774. DOI: 10.3389/fpls.2018.01774.
Lehmann, Ch. O. (1981): Collecting European landraces and development of European gene banks – historical remarks. In: Die Kulturpflanze 29 (1), S. 29–40. DOI: 10.1007/BF02014732.
Miller, J. C.; Tanksley, S. D. (1990): RFLP analysis of phylogenetic relationships and genetic variation in the genus Lycopersicon. In: TAG. Theoretical and applied genetics 80 (4), S. 437–448. DOI: 10.1007/BF00226743.
Persson, K.; Bothmer, R. von (2000): Assessing the allozyme variation in cultivars and Swedish landraces of rye (Secale cereale L.). In: Hereditas 132 (1), S. 7–17. DOI: 10.1111/j.1601-5223.2000.00007.x.
Pons, C.; Casals, J.; Palombieri, S.; Fontanet, L.; Riccini, A.; Rambla, J. L. et al. (2022): Atlas of phenotypic, genotypic and geographical diversity present in the European traditional tomato. In: Horticulture research 9, uhac112. DOI: 10.1093/hr/uhac112.
Rodríguez, G. R.; Muños, St.; Anderson, C.; Sim, S.-Ch.; Michel, A.; Causse, M. et al. (2011): Distribution of SUN, OVATE, LC, and FAS in the tomato germplasm and the relationship to fruit shape diversity. In: Plant physiology 156 (1), S. 275–285. DOI: 10.1104/pp.110.167577.
Stansell, Z.; Björkman, Th. (2020): From landrace to modern hybrid broccoli: the genomic and morphological domestication syndrome within a diverse B. oleracea collection. In: Horticulture research 7, S. 159. DOI: 10.1038/s41438-020-00375-0.
Tanksley, S. D.; McCouch, S. R. (1997): Seed banks and molecular maps: unlocking genetic potential from the wild. In: Science (New York, N.Y.) 277 (5329), S. 1063–1066. DOI: 10.1126/science.277.5329.1063.
Zeven, A. C. (1998): Landraces: A review of definitions and classification. In: Euphytica 104 (2), S. 127–139. DOI: 10.1023/A:1018683119237.
Zeven, A. C. (2000): Traditional maintenance breeding of landraces: 1. Data by crop. In: Euphytica 116 (1), S. 65–85. DOI: 10.1023/A:1004089816030.
Zeven, A. C. (2002): Traditional maintenance breeding of landraces: 2. Practical and theoretical considerations on maintenance of variation of landraces by farmers and gardeners. In: Euphytica 123 (2), S. 147–158. DOI: 10.1023/A:1014940623838.